


Introduction
Many simple forms of heritage follow the Torahs of Mendel. Dominant traits will ever be expressed when nowadays, and recessionary traits will merely be expressed when two recessionary allelomorphs are present. When traversing a pure homozygous dominant trait with a pure homozygous recessionary trait as the P coevals, it is expected that all the progeny in the F1 coevals will show the dominant trait, since every progeny will have one transcript of the dominant allelomorph from one parent and one transcript of the recessionary allelomorph from the other. In the F2 coevals, the expected result will be a 3:1 phenotypic ratio of dominant to recessive, and a 1:2:1 genotypic ratio of homozygous dominant to heterozygous to homozygous recessive ( Campbell et al. 268 ) .
This simple heritage form explains many of the heritage phenomena exhibited in nature, but some heritage forms travel beyond Mendel’s Torahs of genetic sciences. In uncomplete laterality, neither allelomorph is dominant over the other so the result is a blend of both traits. In codominance, both traits are expressed individually.
In mitochondrial heritage, all progeny will have specific cistrons from the female parent. In X-linked recessionary traits, the allelomorphs are located on the X chromosome, and these conditions often appear in males because they merely have one transcript of the X chromosome ( “Inheritance Patterns” ) . When the exact heritage form is unknown in a cross, the ratios of each type of offspring aid to find if the heritage form follows Mendel’s Torahs or if it is one of the above assortments.
In fruit flies, the ruddy and brown cistrons for oculus colour are located on somatic chromosomes. However, a mutant on thewhitecistron in fruit flies on the X chromosome prevents any oculus colour from developing at all ( “The Genetics of Eye Color” ) . The cistron for white oculus colour is epistatic to the ruddy and white oculus cistrons. This is how fruit flies are able to hold three different oculus colourss – when the white mutant is non present, there will be a simple heritage form between ruddy and sepia eyes. When the mutant is present, the ruddy or sepia eyes will non be expressed because they will be masked behind the white mutant.
Drosophila melanogasterwere used in this process because they reproduce really rapidly and are easy manageable. All their nutrient and H2O demands are taken attention of by the substance called media in the underside of the phial. They are a convenient size because they are non excessively large, but they are little plenty to easy separate traits under a microscope ( “The Fruit Fly and Genetics” ) . The life rhythm of the flies begins as eggs. From the eggs emerge the larvae, which look like bantam worms. The larvae grow through three phases until they reach the pupal phase. The pupae mature and darken in colour for three to four yearss until they break forth from the pupal instance to go grownup flies ( “Development” ) .
In this experiment, three crosses were performed between different assortments of the fruit flyDrosophila Melanogaster. Cross 1 was between a reddish brown eyed female and a wild type male, Cross 2 was between a white eyed female and a wild type male, and Cross 3 was between a ruddy eyed, rudimentary winged female with a reddish brown eyed, normal winged male.
In Cross 1, a simple form of Mendel’s Torahs is predicted to be expressed. Wild type flies with ruddy eyes is the dominant phenotype over reddish brown colored eyes. Sepia colored eyes are a consequence from a recessionary cistron, and merely ensue when two sepia-eyed flies mate or when two heterozygous flies mate. Furthermore, reddish brown colored eyes is non dependent on the sex of the fly, so in the instance of this cross all flies in the F1 coevals should hold ruddy eyes, but be bearers for the reddish brown colored oculus trait.
In the F2 coevals when the heterozygous flies mate, the predicted phenotypic ratio will be 3:1, where for every three ruddy eyed flies there would be one reddish brown colored fly. The related genotypic ratio of homozygous dominant to heterozygous to homozygous recessive will be 1:2:1. Our hypothesis for Cross 1 is if there are no mutants and the cross follows Mendel’s Torahs of independent mixture, so the ratio of ruddy to sepia eyed flies will be 3:1 for the F2 coevals.
In cross 2, sex linked heritage plays a function. The mutant for white coloured eyes is X-linked recessive. When the white eyed female is crossed with a ruddy eyed male, all the males in the F1 coevals should exhibit the mutant, and all the females should hold ruddy eyes. This is because the males can merely accept a recessionary allelomorph from the female parent and the Y chromosome from the male parent which does non transport the mutant for white oculus colour.
The females will have the ruddy cistron from the father’s X chromosome which will cover the white cistron from the female parent. The F2 coevals produced by the white eyed male and heterozygous female will therefore hold a genotypic ratio of 1:1:1:1. Therefore, our hypothesis for Cross 2 for the F2 coevals is that if the cistron for the white oculus mutant is located on the X chromosome, so the phenotypic ratio will be 1:1:1:1 if sex is considered.
In cross 3, the focal point shifted from merely looking at oculus colourss to looking at oculus colourss and flying type. The fruit flies could either hold normal wings or exhibit rudimentary wings, which are shortened. Fliess with rudimentary wings have a defect in their rudimentary cistron located on the 2nd chromosome. So, a dihybrid cross will be used to find the predicted phenotypes and genotypes of the F1 and F2 coevals.
A dihybrid cross uses two traits with two allelomorphs each, and so two different facets of an being are crossed. Vestigial wings are a recessionary trait, so two recessionary allelomorphs must be inherited in order to show the trait. This is besides the instance with reddish brown colored eyes. So when a parent coevals of a ruddy eyed vestigial winged female is crossed with a sepia eyed normal winged male, all of the progeny in the F1 coevals should hold ruddy eyes and normal wings.
The F2 coevals, nevertheless, are produced by heterozygotes and therefore four phenotypes should be seen: ruddy eyed normal, sepia normal, sepia vestigial, and ruddy eyed vestigial. Therefore, our hypothesis for the F2 coevals in Cross 3 is that if both traits follow Mendel’s Torahs of independent mixture for the dihybrid cross, so the predicted phenotypic ratio will be 9:3:3:1.
Methodology
Materials used include:
- Phials
- Microscopes
- Fly sleep and anaesthesia wand
- Paint coppices
- Fly nutrient
- Fly sacking
- Cotton stoppers
- Plain white index card
Procedure:
- First, prepare phials for the fruit flies to populate in. Obtain three glass phials, and gauge a few centimetres of Carolina InstantDrosophilaMedium in each. After, put a few beads of H2O in the civilization and allow it sit a few proceedingss to soak in the medium. At this point, besides put in a fly cyberspace. Obtain F1 flies from teacher for the three crosses.
- Check the phials for life. The flies need to be alive for active genteelness intents. However, guarantee that there are no F2 larvae yet, as this could be misdirecting for the consequences. Check the nutrient for wet, and add H2O with a pipet if the nutrient gets excessively dry.
- Anesthetize the fruit flies. Put the phials of flies upside down in the icebox, as this forces the flies into a province of inaction. After about 10 proceedingss, take the phials out and transport each of the three tubings ( for the separate three crosses ) into three different phials. Mark the three phials with tape for either cross 1, cross 2, or cross 3. Tap the flies into the new phials, and shut it with a cotton stopper with an anaesthesia wand connected to it soaked in Flynap. Wait a few proceedingss for the flies to halt traveling or winging to get down the following process.
- Shake the fruit flies onto a white index card and topographic point the card under the dissecting microscope. Use the paintbrush to travel the flies to the centre of the sing field in order to arouse them and see them for the coveted traits. Record the informations in the information tabular arraies. Males have a solid black venters and sex combs on their forelegs, while females have a striped venters and no sex combs. Additionally, females are by and large larger than males [ 1 ] .
- After entering the information for the flies, place them in the mortuary. Record F1 informations for three yearss, or until F2 larvae are seen.
- Repeat the above process utilizing the same phials, but this clip utilizing merely F2 flies. Record information for three yearss.
- Set the excess flies free, and clean out all the phials exhaustively.
Consequences
Cross 1
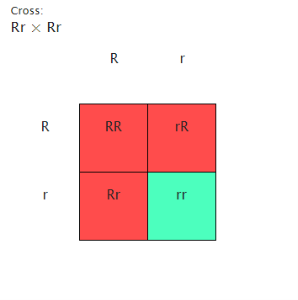
Punnett square: Cross between heterozygous male and heterozygous female for oculus colour
Table 1: Lab Group informations for Cross 1
 Fasdfasdasdffasdfasdf Fasdfasdasdffasdfasdf |
Day 1 | Day 2 | Day 3 | Entire |
| Red M | 3 | 2 | 55 | 60 |
| Red F | 4 | 1 | 58 | 63 |
| Sepia M | 0 | 0 | 6 | 6 |
| Sepia F | 0 | 0 | 4 | 4 |
 Fasdfasdasdffasdfasdf Fasdfasdasdffasdfasdf |
Day 1 | Day 2 | Day 3 | Entire |
| Red M | 2 | 3 | 1 | 6 |
| Red F | 3 | 2 | 0 | 5 |
| Sepia M | 0 | 0 | 0 | 0 |
| Sepia F | 2 | 0 | 0 | 2 |
Table 2: AP Bio 2015 Class Data for Cross 1
 Fasdfasdasdffasdfasdf Fasdfasdasdffasdfasdf |
Pd 8 | Pd 6 | Entire |
| Red M | 39 | 58 | 97 |
| Red F | 46 | 64 | 110 |
| Sepia M | 12 | 7 | 19 |
| Sepia F | 11 | 4 | 15 |
| F2 | sex | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | entire |
| Red M | Meter | 19 | 13 | 8 | 35 | 44 | 21 | 6 | 8 | 36 | 48 | 22 | 260 |
| Red F | F | 37 | 12 | 12 | 38 | 51 | 36 | 5 | 4 | 36 | 50 | 29 | 310 |
| SEPIA M | Meter | 4 | 3 | 5 | 12 | 10 | 6 | 0 | 3 | 5 | 12 | 6 | 66 |
| SEPIA F | F | 5 | 5 | 3 | 16 | 13 | 7 | 2 | 1 | 13 | 14 | 9 | 88 |
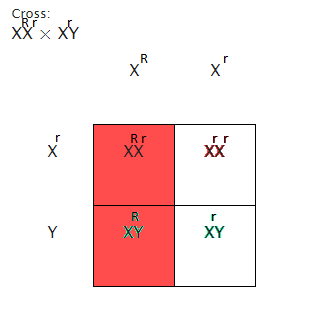
Table 3: Lab Group Data for Cross 2
Expected Sums:
Red M: 1/4 ten x/45 = 11.25
( 11-11.25 ) ^2/11.25 = .006
Red F: 1/4 ten x/45 = 11.25
( 16-11.25 ) ^2/11.25 = 2.01
White M: 1/4 ten x/45 = 11.25
( 8-11.25 ) ^2/11.25 = .939
White F: 1/4 ten x/45 = 11.25
( 10-11.25 ) ^2/11.25 = .139
Expected Sums:
Red M: 1/4 ten x/780 = 195
( 211-195 ) ^2/195 = 1.312
Red F: 1/4 ten x/780 = 195
( 231-195 ) ^2/195 = 6.646
White M: 1/4 ten x/780 = 195
( 162-195 ) ^2/195 = 5.585
White F: 1/4 ten x/780 = 195
( 176-195 ) ^2/195 = 1.851
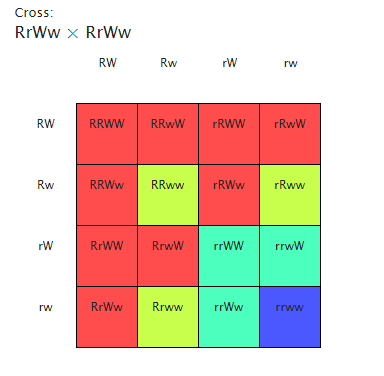
Table 5: Lab Group Data for Cross 3
| F1 | sex | twenty-four hours 2 | twenty-four hours 3 | entire | |||
| Red / NORMAL | Meter | 6 | 16 | 40 | |||
| Red / NORMAL | F | 12 | 17 | 62 | |||
| Red / VESTIGAL | Meter | 0 | 0 | 0 | |||
| Red / VESTIGAL | F | 0 | 0 | 0 | |||
| SEPIA / NORMAL | Meter | 0 | 5 | 5 | |||
| SEPIA / NORMAL | F | 0 | 1 | 1 | |||
| SEPIA / VESTIGAL | Meter | 0 | 0 | 0 | |||
| SEPIA / VESTIGAL | F | 0 | 0 | 0 | |||
| F2 | sex | twenty-four hours 1 | twenty-four hours 2 | twenty-four hours 3 | |||
| Red / NORMAL | Meter | 12 | 7 | 2 | |||
| Red / NORMAL | F | 13 | 13 | 10 | |||
| Red / VESTIGAL | Meter | 1 | 2 | 0 | |||
| Red / VESTIGAL | F | 3 | 3 | 1 | |||
| SEPIA / NORMAL | Meter | 8 | 4 | 1 | |||
| SEPIA / NORMAL | F | 4 | 3 | 4 | |||
| SEPIA / VESTIGAL | Meter | 0 | 0 | 0 | |||
| SEPIA / VESTIGAL | F | 1 | 0 | 1 | |||
Expected Sums:
Red normal: 9/16 ten x/93 = 52.313
( 57-52.313 ) ^2/52.313 = .420
Sepia normal: 3/16 ten x/93 = 17.438
( 24-17.438 ) ^2/17.438 = 2.470
Red vestigial: 3/16 ten x/93 = 17.438
( 10-17.438 ) ^2/17.438 = 3.172
Sepia vestigial: 1/16 ten x/93 = 5.813
( 2-5.813 ) ^2/ ( 5.813 ) = 2.501
Table 6: AP Bio 2015 Group Data for Cross 3
| F1 | sex | 1 | 2 | entire | |||||||||
| Red / NORMAL | Meter | 28 | 37 | 65 | |||||||||
| Red / NORMAL | F | 42 | 51 | 93 | |||||||||
| Red / VESTIGAL | Meter | 5 | 0 | 5 | |||||||||
| Red / VESTIGAL | F | 3 | 0 | 3 | |||||||||
| SEPIA / NORMAL | Meter | 2 | 11 | 13 | |||||||||
| SEPIA / NORMAL | F | 4 | 5 | 9 | |||||||||
| SEPIA / VESTIGAL | Meter | 0 | 0 | 0 | |||||||||
| SEPIA / VESTIGAL | F | 0 | 0 | 0 | |||||||||
| F2 | sex | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | entire |
| Red / NORMAL | Meter | 7 | 14 | 29 | 30 | 16 | 18 | 21 | 26 | 16 | 7 | 10 | 194 |
| Red / NORMAL | F | 21 | 18 | 44 | 34 | 16 | 22 | 36 | 30 | 16 | 11 | 6 | 254 |
| Red / VESTIGAL | Meter | 4 | 3 | 8 | 11 | 4 | 9 | 3 | 0 | 8 | 6 | 11 | 67 |
| Red / VESTIGAL | F | 7 | 8 | 9 | 9 | 3 | 16 | 7 | 0 | 41 | 4 | 16 | 120 |
| SEPIA / NORMAL | Meter | 5 | 4 | 8 | 12 | 7 | 5 | 13 | 4 | 0 | 3 | 3 | 64 |
| SEPIA / NORMAL | F | 5 | 5 | 12 | 14 | 4 | 7 | 11 | 4 | 3 | 2 | 3 | 70 |
| SEPIA / VESTIGAL | Meter | 0 | 1 | 5 | 3 | 1 | 1 | 0 | 0 | 0 | 2 | 2 | 15 |
| SEPIA / VESTIGAL | F | 4 | 2 | 2 | 2 | 2 | 3 | 2 | 0 | 1 | 1 | 12 | 31 |
Expected Sums:
Red normal: 9/16 ten x/815 = 458.438
( 448-458.438 ) ^2/458.438 = .238
Sepia normal: 3/16 ten x/815 = 152.813
( 134-152.813 ) ^2/152.813 = 2.316
Red vestigial: 3/16 ten x/815 = 152.813
( 187-152.813 ) ^2/152.813 = 7.648
Sepia vestigial: 1/16 ten x/815 = 50.938
( 46-50.938 ) ^2/50.938 = 0.479
Discussion of Results and Decisions
The chi-square statistical analysis trials calculated the undermentioned consequences for the lab group informations: accept cross 1, accept cross 2, and reject cross 3. The trials calculated the following for the AP Bio Class information: cull cross 1, reject cross 2, reject cross 3. Therefore, our first two void hypotheses were supported for our little group informations while the tierce was rejected, and the void hypotheses were all rejected by the category information. To reject a void hypothesis does non intend that the hypothesis is so wrong. Rejecting a void hypothesis means that the sample observ
ed demonstrated a important difference from the anticipations of the void hypothesis, plenty to reason that the void hypothesis is false. However, that does non intend it really is false. For the sample examined in the process, the void hypothesis did non fulfill the consequences, but this could be due to opportunity events and non a defect in the theory.
In Cross 1, the F1 coevals displayed the simple heritage form as predicted by Mendel’s Torahs, until twenty-four hours 3 was recorded. On the 3rd twenty-four hours, reddish brown eyed males and females were seen. This is because information was collected after the F2 pupa hatched. Once the F2 flies were added to the mix, consequences began to look that were non predicted by our hypothesis. However, this does non intend the hypothesis was wrong ; instead, the informations aggregation was contaminated by the presence of the F2.
The simple manner to avoid this job would be to invalidate any informations collected one time F2 larvae Begin to look. If the information had been collected sooner, with less of a spread between aggregation, so the F1 flies would non hold had clip to reproduce, and the informations would be pure. This besides happened in Cross 2 and in Cross 3, but merely on the 3rd twenty-four hours. Otherwise, the F1 informations supported the hypotheses for all crosses.
For the simple forms of heritage, all the F1 flies were heterozygous for their traits, and no recessionary traits were exhibited. For the sex-linked heritage in Cross 2, all males were hemizygous white because they inherited merely one X chromosome, which was from the white eyed female parent. The females were all ruddy eyed because they received the dominant ruddy allelomorph from the male parent.
All the informations, whether it accepted or rejected the void hypothesis, supported the averments made in the debut that ruddy eyes are dominant to sepia eyes, and that white eyes are recessionary. The information besides supported that rudimentary wings are a recessionary trait. The mistake with the void hypothesis comes into drama when the ratios of the specific ascertained traits are compared to the expected ratios.
The lab group informations from cross 1 and traverse 2 supports the anticipations made by Mendel’s Torahs. The predicted result ratio of the first cross was 3:1, and in the experiment the ratio was 11:2. Harmonizing to chi-square statistical analysis, this ratio was near adequate to the predicted result that it confirmed the void hypothesis. For the 2nd cross, the predicted result was 1:1:1:1, following sex-linked recessionary heritage forms.
The recorded experimental ratio was 11:16:8:10, which one time once more was close plenty for the chi-square trial to accept the void hypothesis. As for cross 3, although the divergence was extremely important, that does non intend that the void hypothesis was wrong, as mentioned antecedently. The hypothesis did so foretell all the phenotypes that were expressed in the F2 flies, so opportunity events could hold skewed the ratios.
One possible beginning of mistake is that the flies could hold been falsely sexed or identified. Without paying close attending to the sex combs or abdominal forms, the fly could be falsely labeled. Additionally, the pigment coppice could hold crushed the wings of the flies, giving them a rudimentary visual aspect. Both errors would change the ratio observed. Following, there could hold been mutants in the flies that altered the consequences.
These are wholly random events, so they could hold happened in one of the two traits, or both, or none. Additionally, by chance killing flies, acquiring them stuck in the nutrient, or by chance cathartic flies would wholly be opportunity ways to change the ratios. Despite any beginnings of mistake, the process was still a valuable manner to detect heritage forms in nature.
Bibliography
- Campbell, Neil, et Al. Chapter 15: The Chromosomal Basis of Inheritance. ”Biology: AP Edition. By Neil A. Campbell. 8th erectile dysfunction. San Francisco: Pearson Education, 2008. 286-92. Print.
- “Development of the Fruit Fly Drosophila melanogaster.” Place of Publication Not Identified: Stationery Office, 2006. Web. 6 June 2015.
- The Fruit Fly and Genetics. ”The Fruit Fly and Genetics. N.p. , n.d. Web. 06 June 2015.
- The Genetics of Eye Colour. ”The Arrogant Scientist. N.p. , 29 Jan. 2009. Web. 31 May 2015.
