



Gene Alteration:
Inroduction: The construct of cistron has evolved and go more complex since it was foremost proposed. The term Gene was foremost proposed by Wilhelm Johannsen. Gene is a functional section of bases in familial stuff. It means that the functional cistron transcribed into the functional merchandises like RNAs, proteins etc. Each cistron may be in alternate signifiers called allelomorphs, which have codification for different familial character. Alleles of cistron are busying matching places on homologous chromosomes and commanding the same features but bring forthing different effects.
If any alterations in homologous allelomorphs or complete cistron take topographic point due to some internal every bit good as external effects causes change of cistron besides known mutant. Mutants can take topographic point through cistron loss, elaboration, rearrangement etc. After mutant, the merchandise synthesised by written text can be good or bad for the being. Sometimes, it can take to many diseases.
Mutant:
Genome is non a inactive entity. It is capable to different types of heritable alterations. A sudden and heritable alteration in the sequences of an organisms genome that give rise to jump signifiers of any cistron is called mutant. An disconnected alteration in a genotype of an being that is non the consequence of recombination, is considered as mutant. The procedure by which mutants is produced is known mutagenesis, which lead to formation of mutant being. Broadly talking, mutants include all type of heritable familial alteration of an being. This type of familial alterations include chromosomal aberrances and alterations in single cistrons and ploidy. By and large, the alterations in the cistron is known as cistron mutant.
General features of mutant:
- By and large recessionary mutants occur but dominant mutants besides occur.
- Mutants are by and large harmful to the beings.
- Mutants are random, occur at any clip and in any cell of an being.
- Mutants are perennial i.e. the same mutant may happen once more and once more.
Role of mutant:
- Ultimate beginning of all familial fluctuation and it provides the natural stuff for your development.
- Mutant consequences into the formation of allelomorphs. Without mutant, all cistrons would be in merely one signifier.
- Organisms would able to germinate and accommodate to environmental alteration.
Molecular footing of Gene Mutation:
Mutants arise in two ways:
- Spontaneous mutant
- Induced mutant
Spontaneous mutant: Some mutants are self-generated that history without intervention of the being with an exogenic mutagen. Mutagens are the agent which increase the frequence of happening of mutants. Spontaneous mutants chiefly account for the back-ground rate of mutant and are presumptively the ultimate beginning of natural familial fluctuation in populations. Spontaneous mutants can happen because of reproduction mistakes & A ; self-generated lesions & A ; heterotaxy of permutable elements during their normal growing of the cell.
Induced mutants: These mutants arise because a mutagen has reacted with the parent DNA, doing a structural alteration that affects the base-pairing capableness of the altered bases.
Mistake in DNA reproduction:
Each of common bases in DNA can spontaneously undergo a transeunt rearrangement of portending. A proton switching within nitrogen-bearing base forms one of its rare tautomeric signifier ( tautomeric displacement ) . Formation of the tautomer of any basal alters its base-pairing belongingss.
However, if a base existed in the rare signifier at the minute that it was being replicated or being incorporated into a nascent Deoxyribonucleic acid concatenation, a mutant might ensue. When the bases are present in their rare imimo or enol provinces, they can organize adenine-cytosine and guanine-thymine base brace. The net consequence of such type event & A ; the subsequent reproduction segregate the mismatched base brace, is an AT to GC or a GC to AT base brace permutation.
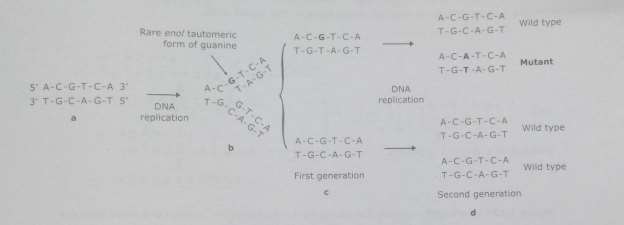
Fig. Mutation via tautomeric displacements in bases of DNA.
Mistake during DNA reproduction may ensue in permutation mutant ( point mutant ) or frameshift mutant.
Substitution mutant: A cistron mutant that consequences from the permutation of one base brace for another is known as point mutant. It may be passage or trans-version mutant.
Passage: Passage, the most common category, consisting the permutation of one pyrimidine by other, or of one purine by the other nucleosides. In passage, a G-C brace is replaced with an A-T brace or frailty versa.
Transversion: If purine is replaced by pyrimidine, or vice-versa so it is known as transversion.
Most of mispairing mutants are passages. It is because of an AC or GT mispair does non falsify the DNA dual spiral every bit much as AG or CT base brace distort. However, transversions besides can take topographic point through mispairing.
Frame-shift Mutant: Aberrant reproduction can besides ensue in little figure of extra-nucleotides being inserted into the polynucleotide being synthesized, and/or some bases in the templet non being copied. Addition or omission of base braces that occurs within the protein-coding part of a cistron have the consequence of switching the translational reading frame. In bulk of the instances, these consequences in a failure to synthesise a functional protein, therefore, leting the mutant to be identified by its phenotypic effects. Because these mutants cause a displacement in the translational reading frame, so such mutants are known as frame-shift mutants. Frame-shift mutants are peculiarly more prevailing when the templet Deoxyribonucleic acid contains short repeated sequences. This is because perennial sequences can bring on the reproduction slippage, in which the templet strand and its transcript switch their comparative place so that the way of templet is either copied twice or missed out. The consequence is that the new polynucleotide has a larger or smaller figure, severally, of the repeated units.
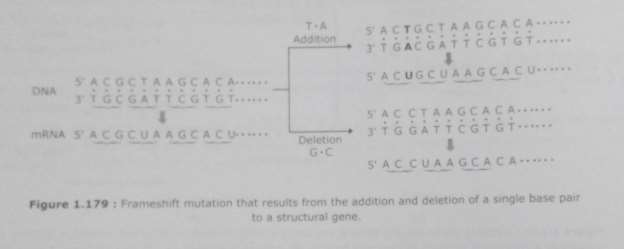
Many nucleotide repeat enlargement disease in human occur as a consequence of reproduction slippage. For illustration, the human HD cistron contains the sequence 5’-CAG-3’ repeated between 6 and 35 times, holding codification for a series of glutamines in the protein merchandise. In the instance of Huntington’s disease, this repetition expands between 36-121 which increasing the length of the polyglutamine piece of land and ensuing in a dysfunctional protein.
Spontaneous lesions: Naturally happening harm to the Deoxyribonucleic acid, called spontaneously lesions, besides can take mutants. Depurination and deaminization are most frequent type of self-generated lesions.
Deamination: It is the remotion of an amino group from a molecule. 3 of the 4 nitrogen-bearing bases usually present in DNA ( C, A, G ) contain exocyclic amino group. Deamination of C consequences in the formation of U. Similarly deaminization of A and G consequence in the formation of hypoxanthine and xanthine, severally. Deamination changes the standard base coupling forms.
Depurination and Depyrimidination: Depurination is the loss of purine due to interrupting of glycosidic bond of bases in DNA. Similarly, depyrimidination is the loss of pyrimidine base. Depurination and depyrimidination are more common at acidic pH. However, these procedures can besides happen at appreciable rates at nuetral or alkalic pH. Pyrimidine nucleosides are well more stable than purine nucleosides with regard to the glycosidic bond of the bases to deoxyribose. The mechanism of depyrimidination is the same as for depurination.
Transposition: A jumping gene is a Deoxyribonucleic acid sequence able to infix itself at a new location in the genome without holding any sequence relationship with the mark venue. Interpolation of a movable elements into or near a functional cistron can change its look by doing loss of cistron map or by altering the gene’s look ( insertion mutant ) .
Loss and addition of map mutants:
In principle mutant of a cistron might do a phenotypic alteration in either of two ways:
- Loss of map ( void ) mutant: the synthesized protein or RNA ( merchandise ) may hold reduced or no map.
- Addition of map mutant: the synthesized protein or RNA ( merchandise ) may hold increased or new map.
Most of the clip there is a loss of map in the synthesized merchandise due to the debut of random familial alterations because of mutants. By and large, ‘loss of map mutations’ are found to be recessionary. In a diploid wild type cell, there are 2 wild type allelomorphs of a cistron, both doing normal cistron merchandise. In heterozygote, the individual wild type allelomorph may be plenty for the formation of normal cistron merchandises to bring forth a wild type phenotype. In such instances, ‘loss of map mutations’ are recessionary. However, some ‘loss of map mutations’ are dominant. In these instances, the individual wild type allelomorph in the heterozygote can non supply the formation of adequate cistron merchandise needed for the cell to be wild type. ‘Gain of map mutations’ normally cause dominant phenotypes as the presence of a normal allelomorph does non forestall the mutant allelomorph from acting abnormally.
Chromosome Aberrances:
Omission: A omission is an aberrance, in which there is a loss of a section of a chromosome.
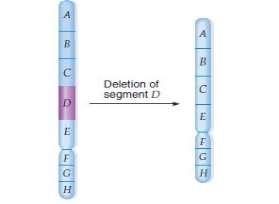
Omission can be of two types- terminus and interstitial. Terminal omission involves a individual interruption whereas in interstitial omission two interruptions are required. A omission of any part of a chromosome is normally deadly to a diploid being because it causes familial instability. When an being heterozygous for a brace of allelomorphs, A and a, loses a little part of the chromosome bearing the dominant allelomorph, the recessionary allelomorph on the other chromosome will go uttered phenotypically. This is called pseudodominance.
Duplicate: A duplicate is the happening of a chromosome in two or more transcripts per genome.
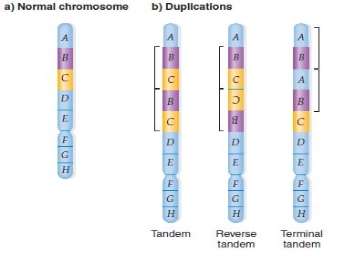
Duplicate can be tendem or contrary. The duplicate parts can be located next to each other, or dispersed on the same chromosome.
Inversions:If two interruptions in one chromosome and the part between the interruptions rotates 180 grades before rejoining with the two terminal fragments, so such an event creates a chromosomal mutant called an inversion.
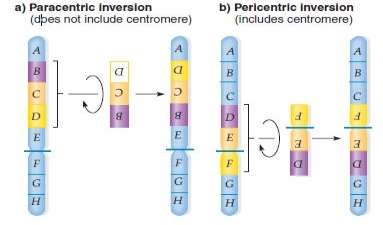
Unlike omissions and duplicates, inversions do non alter the overall sum of the familial stuff, so inversions are by and large feasible and demo no peculiar abnormalcies at the phenotypic degree. In some instances, one of the chromosome brakes is within a cistron of indispensable map, and so that breakpoint Acts of the Apostless as a deadly cistron mutant linked to the inversion.
The location of the centromere relation to the inverted section determinants the familial behavior of the chromosome. If the kinetochore is outside the inversion, so the inversion is said to be para-centric, whereas inversions crossing the centromere pericentric.
Translocations: When two non-homologous chromosomes exchange their parts, the ensuing chromosome rearrangements are translocations. Translocation may be mutual or non-reciprocal.
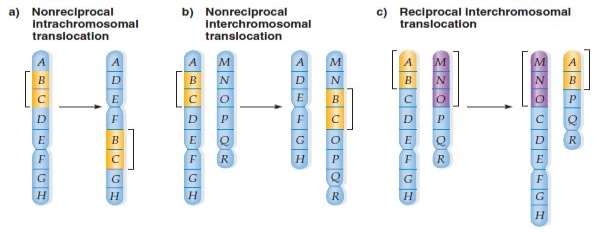
Non-reciprocal translocation involves the transportation of a section in one way from one chromosome to another. In mutual translocations, the most common type- a section from one chromosome is exchanged with a section from another non-homologous one, so two translocation chromosomes are generated at the same time. The exchange of chromosome parts between non-homologs establishes new linkage dealingss.
Applications of Gene Changes:
Site directed mutagenesis:
Site directed mutagenesis techniques are used to change the nucleotide sequence of the cistron coding for a protein of involvement. Conventional mutagenesis is a random procedure that introduces alterations at unspecified places DNA molecule. Hence showing of big Numberss of mutated beings is necessary to happen a mutant of involvement.
A assortment of DNA uses can be used to present mutants into cloned cistrons. The simplest is to cancel a limitation fragment, to open the Deoxyribonucleic acid at a limitation site and take few bases, or to infix new DNA section at a limitation site ( termed cassette mutagenesis ) . These are comparatively big graduated table alterations. By the technique called oligonucleotide directed mutagenesis, one can change a individual base at specified place. There are a figure of different ways of transporting out oligonucleotide directed mutagenesis.
One version of oligonucleotides directed mutagenesis is described here. First, a single-stranded version of the cistron is obtained by cloning in M13 vector. A short oligonucleotide is so synthesized, complementary to the relevant part of the cistron, but incorporating the desired nucleotide change. This base is hybridized to the Deoxyribonucleic acid and used as the primer for a strand synthesis reaction that is allowed to go on all the manner around the round templet molecule.
The freshly formed heteroduplex is used to transform cells. After debut into E. Coli, DNA reproduction produces legion transcripts of this recombination DNA molecule. Half the ensuing double-stranded molecules are transcripts of the original strand of DNA, and the staying are transcripts of the strand that contains the mutated sequence. The double-stranded molecules direct synthesis of phage atoms, so about half the phages released from the septic bacterium carry a single-stranded version of the mutated molecule. The phages are plated on to solid agar so that plaques are produced, and the mutant 1s identified by hybridisation examining with the original oligonucleotide. The mutated cistron can so be placed back in its original host by homologus recombination or transferred to an E. coli vector designed for the synthesis of protein from cloned DNA, so that a sample of mutated protein can be obtained.
Gene therapy:
Gene therapy is a technique for rectifying faulty cistrons responsible for disease development. Gene therapy typically aims to supplement a faulty mutation allelomorph with a functional 1.
Scientist may utilize one of several attacks for rectifying faulty or unnatural cistrons:
- A normal cistron may be inserted into a non-specific location within the genome ( cistron add-on ) . This is the most common attack.
- An unnatural cistron can be replaced by a normal cistron through homologus recombination ( cistron replacing ) .
- An unnatural cistron can be repaired through selective contrary mutant, which returns the cistron to its normal map.
Gene therapy may be germ-line and bodily cell cistron therapy. Current cistron therapy is entirely bodily cistron therapy which involves the debut of cistrons into bodily cells of an affected person. Germ-line cistron therapy involves the lasting catching alteration of the genome of a gamete, a fertilized ovum or an early embryo.
Potential usage of bodily cistron therapy:
The possible usage of this is to bring around familial diseases. The first instance of cistron therapy in 1990, a 4 twelvemonth old patient with combined immunodeficiency received an extract of white blood cells that had been genetically modified to incorporate the cistron that was non functional in his genome. Since so, cistron therapy has been studied and by experimentation tested for several medical conditions like Hemophilia A, Hemophilia B, Cystic fibrosis, Cancer etc.
Mentions:
Books:
- iGenetics A molecular Approach, 3rdedition by Peter J. Russell.
- Molecular Biology of The Cell 5Thursdayedition by Prince alberts
- Molecular Biology of The Gene by Watson and Bell
